Aging is part of normal human development. It is define as the
accumulation of random changes in cells and tissues (Harman D., 2001). Unfortunately,
the majority of random changes increase the chances of developing diseases and death.
The average life span of humans in developed countries has slowly increased due to
improvements in living conditions and advances in medical technology. However, neither
could reduce the rate of aging. Hence, the maximum life span of humans has remained the same.
One theory on the rate of aging is that it correlates with the rate of molecular damage
accumulation in cells and tissues. Reactive Oxygen Species (ROS) produced in the course
of normal metabolism causes molecular damage in cells (Hekimi et al., 2001). Once the limit
of molecular damage an organism can sustain is reached, death occurs. Experiments that
demonstrate reducing ROS damage extends life span supports this theory (e.g., Felkai et al.,
1999; and Hekimi et al., 2001).
Role of C. elegans in the study of aging
Studying gene mutations in C. elegans that extend life span have led to
a better understanding in aging genetics. These studies have the potential to develop methods
to reduce the rate of aging in humans. Gerontogenes are genes that when altered, cause life
extension in the organism (Harman D., 2001). C. elegans have two main classes of gerontogenes.
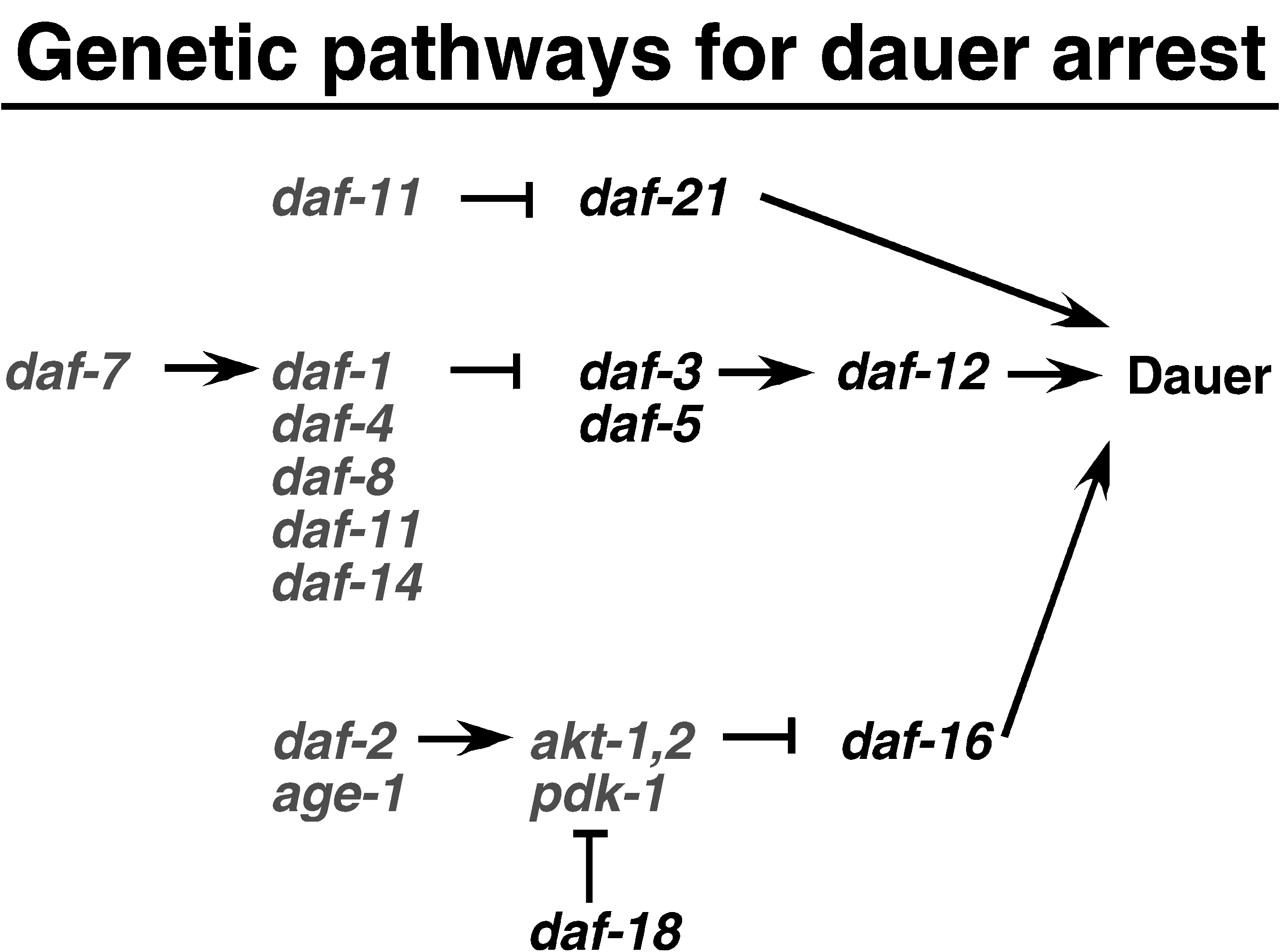
Genes in the first class are involved in the regulation of C. elegans diapause (Finch et al., 2001).
C. elegans can either arrest at the dauer diapause stage or grow reproductively. Under adverse
conditions, such as food shortage, and/or high environmental temperature, developing into dauer
larvae are favoured because they are more stress resistant (Hekimi et al., 2001). DAF-2 signalling
determines which developmental pathway C. elegans takes (Finch et al., 2001). For example, strong
DAF-2 signalling leads to growth and reproduction, whereas absence of DAF-2 signalling leads to
constitutive dauer arrest. C. elegans having reduction function mutations in the daf-2 gene have
longer life span when compared to wild type C. elegans. One possible explanation to the observation
is that the mutations cause an inappropriate expression of dauer developmental program in the
animal. Consequently, stress protective enzymes are upregulated enhancing protection against ROS
(Hekimi et al., 2001). Hence, accumulation of molecular damage in cells is reduced. Reduction
function mutations in other genes in the daf-2 pathway have also been observed to extend life
span, such as age-1 and pdk-1.
In the second class, mutated genes cause life processes of the mutants to be slower than the
wild type C. elegans. These processes include the duration of cell cycle, and embryonic and
postembryonic development. In addition, adult behaviours, such as defecation cycle, swimming
and pharyngeal pumping are lengthened (Hekimi et al., 2001). The clk-1 gene is one of the
well-characterised genes in this class. Clk-1 encodes a protein that is expressed in the mitochondria
of every tissue (Hekimi et al., 2001). Compare to wild type C. elegans, respiration rates in clk-1
mutants are lower. In addition, clk-1 mutants lack UQ9, a component of the electron transport in
the mitochondria. Instead, DMQ9, an intermediate in UQ9 biosynthesis, is present that replaces UQ9
as the electron acceptor in electron transport (Miyadera et al., 2001). Both differences are thought
to extend life span of clk-1 mutants by reducing the rate of ROS production. Other mutated genes in
the same class include clk-2, clk-3 and gro (Hekimi et al., 2001).
To read about a sepecific genomic experiment, click here.
The future of aging studies in C. elegans
Although many gene mutations resulting in extension of life span in C. elegans have
been identified, it is still a mystery how these mutations are correlated with longer
life span. In addition, the pathways mutated genes are involved in remain unknown, such
as the clk-1 gene. Recently, the entire genome of C. elegans has been sequenced and a
gene expression map of C. elegans has been produced (Kim et al., 2001). Related genes
are grouped together in the gene expression map. Therefore, once a gene is located on
the map, then all of the related genes are also revealed. In the case of clk-1, the
procedure could reveal pathways in which clk-1 are involved in. Ultimately, it could
lead to identification of all of clk-1's functions. Aging is a complex process involving
many genes. The gene expression map enables us to identify all the pathways simultaneously,
which was not possible before.
For genes capable of affecting life span in C. elegans with known orthologs in humans, whether
mutations in the human ortholog have the same affect remain to be determined. For example, daf-2
is an ortholog of the human insulin receptor. Could a reduction function mutation in the human
insulin receptor also result in life span extension? Neurons have been hypothesised as a primary
determinant of life span because they may be particularly sensitive to ROS (Finch et al., 2001).
Microarray experiments monitoring gene transcripts of the neurons could identify mRNAs critical
in aging. Afterwards, identification of similar mRNA in mammalian brain could be done using microarrays.
Microarray could also be used to compare aging in different tissues in C. elegans. The procedure could
reveal changes that serve as molecular markers for the aging process. Once it is obtain, identification
of how each mutant affects aging would be a lot easier. The study of aging in C. elegans is just
beginning. One day, researchers might be able to decrease the aging process in humans. People who
are sixty might look and feel as if they are only twenty years old. The maximum life span of a
person could be many times longer than it is possible at present.
references
Diagram courtesy of:
Finch, C., E., Ruvkun, G., 2001. The Genetics of Aging. Annual Review of Genomics and Human Genetics 2:435-62.
Harman, D. 2001. Aging: Overview. Annals of the New York Academy of Sciences 928:1-21.
Hekimi, S., Burgess. J., Bussiere, F. Meng, Y., Benard, C. 2001. Genetics of lifespan in C. elegans: molecular diversity, physiological complexity, mechanistic simplicity. Trends in Genetics 17:712-718.
Kim, S.K., Lund, J., Kiraly, M., Duke, K., Jiang, M., Stuart, J.M., Eizinger, A., Wylie, B.N., Davidson, G.S. 2001. A Gene Expression Map for Caenorhabditis elegans. Science 293:2087-2092.
Miyadera, H., Amino, H., Hiraishi, A., Taka, H., Murayama, K., Miyoshi, H., Sakamoto, K., Ishii, N., Hekimi, S., Kita, K. 2001. Altered Quinone Biosynthesis in the Long-lived clk-1 Mutants of Caenorhabditis elegans. The Journal of Biological Chemistry 276:7713-7716.
Pierce, S., B., Costa, M., Wisotzkey, R., Devadhar, S., Homburger, S., A., Buchman, A., R., Ferguson, K., C., Heller, J., Platt, D., M., Pasquinelli, A., A., Liu, L., X., Doberstein, S., K., Ruvkun, G. 2001. Regulation of DAF-2 receptor signaling by human insulin and ins-1, a member of the unusually large and diverse C. elegans insulin gene family. Genes and Development 15(6):672-686.
Wolkow, C., A., Kimura, K., D., Lee, M., S., Ruvkun, G., 2000. Regulation of C. elegans life-span by insulinlike signaling in the nervous system. Science 290(5489):147-50.